Математическая модель хищник жертва примеры. Равновесие хищник—жертва. по дисциплине «Моделирование систем»
В модели Колмогорова сделано одно существенное допущение: поскольку предполагается, что при то это означает существование в популяции жертв механизмов, регулирующих их численность даже в отсутствие хищников.
К сожалению, подобная формулировка модели не позволяет ответить на вопрос, вокруг которого в последнее время ведется много споров и о котором мы уже упоминали в начале главы: каким образом популяция хищников может оказывать регулирующее воздействие на популяцию жертв с тем, чтобы вся система была устойчива? Поэтому мы вернемся к модели (2.1), в которой механизмы саморегуляции (например, регулирование при помощи внутривидовой конкуренции) в популяции жертв (впрочем, как и в популяции хищников) отсутствуют; поэтому единственным механизмом регуляции численностей, входящих в сообщество видов, являются трофические взаимоотношения между хищниками и жертвами.
Здесь (так что, в отличие от предыдущей модели, Естественно, что решения (2.1) зависят от конкретного вида трофической функции которая, в свою очередь, определяется характером хищничества, т. е. трофической стратегией хищника и защитной стратегией жертвы. Общими для всех этих функций (см. рис. И) являются следующие свойства:
Система (2.1) имеет одну нетривиальную стационарную точку, координаты которой определяются из уравнений
![]()
при естественном ограничении .
Существует еще одна стационарная точка (0, 0), соответствующая тривиальному равновесию. Нетрудно показать, что эта точка - седло, а сепаратрисами являются координатные оси.
Характеристическое уравнение для точки имеет вид
![]()
Очевидно, что для классической вольтерровской модели .
Поэтому величину f можно рассматривать как меру отклонения рассматриваемой модели от вольтерровской.
![]()
стационарная точка - фокус, и в системе появляются колебания; при выполнении противоположного неравенства - узел, и колебания в системе отсутствуют. Устойчивость этого состояния равновесия определяется условием
т. е. существенным образом зависит от вида трофической функции хищника.
Условие (5.5) можно интерпретировать следующим образом: для устойчивости нетривиального равновесия системы хищник - жертва (и тем самым, для существования этой системы) достаточно, чтобы в окрестности этого состояния относительная доля потребляемых хищником жертв возрастала с возрастанием численности жертв. Действительно, доля жертв (от общей их численности), потребляемых хищником, описывается дифференцируемой функцией условие возрастания которой (положительность производной) выглядит как
![]()
Последнее условие, взятое в точке есть не что иное, как условие (5.5) устойчивости равновесия. При непрерывности оно должно выполняться и в некоторой окрестности точки Таким образом, если численности жертв в этой окрестности, то
Пусть теперь трофическая функция V имеет вид, изображенный на рис. 11, а (характерный для беспозвоночных). Можно показать, что для всех конечных значений (поскольку выпукла вверх)
т. е. при любых величинах стационарной численности жертв неравенство (5.5) не выполняется.
Это означает, что в системе с таким видом трофической функции не существует устойчивого нетривиального равновесия. Возможны несколько исходов: либо численности как жертвы, так и хищника неограниченно возрастают, либо (при прохождении траектории вблизи одной из координатных осей) в силу случайных причин численность жертвы или численность хищника станет равной нулю. При гибели жертвы через некоторое время погибнет и хищник, если же сначала погибает хищник, то численность жертвы начнет экспоненциально возрастать. Третий вариант - возникновение устойчивого предельного цикла - невозможен, что легко доказывается.
В самом деле, выражение

в положительном квадранте всегда положительно, если только имеет вид, изображенный на рис. 11, а. Тогда, согласно критерию Дюлака в этой области отсутствуют замкнутые траектории и устойчивого предельного цикла существовать не может.
Итак, можно сделать вывод: если трофическая функция имеет вид, изображенный на рис. 11, а, то хищник не может быть регулятором, обеспечивающим стабильность популяции жертвы и тем самым устойчивость всей системы в целом. Система может быть устойчива только в том случае, когда в популяции жертвы существуют собственные внутренние регулирующие механизмы, например, внутривидовая конкуренция или эпизоотии. Этот вариант регулирования был уже рассмотрен в §§ 3, 4.
Ранее отмечалось, что такой тип трофической функции характерен для хищников-насекомых, «жертвами» которых также обычно являются насекомые. С другой стороны, наблюдения за динамикой многих природных сообществ типа «хищник - жертва», включающих в себя виды насекомых, показывают, что для них характерны колебания очень большой амплитуды и весьма специфического вида.
Обычно после более или менее постепенного роста численности (который может происходить либо монотонно, либо в виде колебаний с возрастающей амплитудой) происходит ее резкое падение (рис. 14), и затем картина повторяется. По-видимому, такой характер динамики численностей видов насекомых может быть объяснен неустойчивостью этой системы при малых и средних значениях численности и действием мощных внутрипопуляционных регуляторов численности при больших ее значениях.

Рис. 14. Динамика численности популяции австралийской листоблошки Cardiaspina albitextura, кормящейся на эвкалиптах. (Из статьи: Clark L. R. The population dynamics of Cardiaspina albitextura.-Austr. J. Zool., 1964, 12, № 3, p. 362-380.)
Если же в систему «хищник - жертва» входят виды, способные к достаточно сложному поведению (например, хищники способны к обучению или жертвы способны находить убежище), то в такой системе возможно существование устойчивого нетривиального равновесия. Доказывается это утверждение весьма просто.
В самом деле, трофическая функция тогда должна иметь вид, изображенной на рис. 11, в. Точка на этом графике - точка касания прямой, проведенной из начала координат, графика трофической функции Очевидно, что в этой точке функция имеет максимум. Также легко показывается, что для всех выполняется условие (5.5). Следовательно, нетривиальное равновесие, в котором численность жертв меньше будет асимптотически устойчиво
Однако мы ничего не можем сказать о том, сколь велика область устойчивости этого равновесия. Например, если существует неустойчивый предельный цикл, то эта область должна лежать внутри цикла. Или другой вариант: нетривиальное равновесие (5.2) неустойчиво, но существует устойчивый предельный цикл; в этом случае также можно говорить о стабильности системы хищник - жертва. Поскольку выражение (5.7) при выборе трофической функции типа рис. 11, в может менять знак при изменении при , то критерий Дюлака здесь не работает и вопрос о существовании предельных циклов остается открытым.

КОМПЬЮТЕРНАЯ МОДЕЛЬ «ХИЩНИК-ЖЕРТВА»
Казачков Игорь Алексеевич 1 , Гусева Елена Николаевна 2
1 Магнитогорский государственный технический университет им. Г.И. Носова, институт строительства, архитектуры и искусства, студент 5 курса
2 Магнитогорский государственный технический университет им. Г.И. Носова, институт энергетики и автоматизированный систем, кандидат педагогических наук, доцент кафедры бизнес-информатики и информационных технологий
Аннотация
Данная статья посвящена обзору компьютерной модели «хищник-жертва». Проведенное исследование позволяет утверждать, что экологическое моделирование играет огромную роль в исследовании окружающей среды. Данная проблематика имеет многогранный характер.
COMPUTER MODEL «PREDATOR-VICTIM»
Kazatchkov Igor Alekseevich 1 , Guseva Elena Nikolaevna 2
1 Nosov Magnitogorsk State Technical University, Civil Engineering, Architecture and Arts Institute, student of the 5th course
2 Nosov Magnitogorsk State Technical University, Power Engineering and Automated Systems Institute, PhD in Pedagogical Science, Associate Professor of the Business Computer Science and Information Technologies Department
Abstract
This article provides an overview of the computer model "predator-victim". The study suggests that environmental simulation plays a huge role in the study of the environment. This problem is multifaceted.
Для исследования окружающей нас среды используют экологическое моделирование. Математические модели используют в тех случаях, когда нет естественной среды и нет естественных объектов, она помогает сделать прогноз влияния разных факторов на исследуемый объект. Данный метод берет на себя функции проверки, построения и интерпретацию полученных результатов. На основе таких форм экологическое моделирование занимается оценкой изменений, окружающей нас среды.
В настоящий момент подобные формы используется для изучения окружающей нас среды, а когда требуется изучить какую-либо из ее областей, то применяют математическое моделирование. Данная модель дает возможность спрогнозировать влияние тех или иных факторов на объект изучения. В свое время был предложен тип «хищник – жертва» такими учеными как: Т. Мальтусом (Malthus 1798, Мальтус 1905), Ферхюльстом (Verhulst 1838), Пирлом (Pearl 1927, 1930), а также А. Лотки (Lotka 1925, 1927) и В. Вольтерры (Volterra 1926).Эти модели воспроизводят периодический колебательный режим, возникающий в результате межвидовых взаимодействий в природе.
Одним из основных методов познания является моделировка. Помимо того, что в нем можно спрогнозировать изменения, происходящие в окружающей среде, к тому же помогает найти оптимальный способ решения проблемы. Уже давно в экологии используют математические модели, для того чтобы установить закономерности, тенденции развития популяций, помогают выделить суть наблюдений. Макет может служить образцом поведения, объекта.
При воссоздании объектов в математической биологии используются прогнозирования различных систем, предусматриваются специальные индивидуальности биосистем: внутренне строение особи, условия жизнеобеспечения, постоянство экологических систем, благодаря которым сберегается жизнедеятельность систем.
Появление компьютерного моделирования значительно раздвинуло рубеж способностей исследования. Возникло вероятность многосторонней реализации трудных форм, не допускающих аналитического изучения, появились новейшие направления, а еще имитационное моделирование.
Рассмотрим, что же такое объект моделирования. «Объектом является замкнутая среда обитания, где происходит взаимодействие двух биологических популяций: хищников и жертв. Процесс роста, вымирания и размножения происходит
непосредственно на поверхности среды обитания. Питание жертв происходит за счет тех ресурсов, которые присутствуют в данной среде, а питание хищников происходит за счет жертв. При этом питательные ресурсы могут быть как возобновляемые, так и не возобновляемые.
В 1931 году Вито Вольтеррой были выведены следующие законы отношения хищник-жертва.
Закон периодического цикла – процесс уничтожения жертвы хищником нередко приводит к периодическим колебаниям численности популяций обоих видов, зависящим только от скорости роста плотоядных и растительноядных, и от исходного соотношения их численности.
Закон сохранения средних величин – средняя численность каждого вида постоянна, независимо от начального уровня, при условии, что специфические скорости увеличения численности популяций, а также эффективность хищничества постоянны.
Закон нарушения средних величин – при сокращении обоих видов пропорционально их числу, средняя численность популяции жертвы растет, а хищников – падает.
Модель хищник-жертва – это особая взаимосвязь хищника с жертвой, в результате которой выигрывают оба. Выживают наиболее здоровые и приспособленные особи к условиям среды обитания, т.е. все это происходит благодаря естественному отбору. В той среде где нет возможности для размножения, хищник рано или поздно уничтожит популяцию жертвы, в последствии чего вымрет и сам» .
На земле существует множество живых организмов, которые при благоприятных условиях увеличивают численность сородичей до огромных масштабов. Такая способность называется: биотический потенциал вида, т.е. увеличение численности вида за определенный промежуток времени. Каждый вид имеет свой биотический потенциал, к примеру крупные виды организмов за год могут возрасти всего в 1,1 раза, в свою очередь организмы более мелких видов, таких как рачки и т.д. могут увеличить свой вид до 1030 раз, ну а бактерии еще в большем количестве. В любом из этих случаев популяция будет расти в геометрической прогрессии.
Экспоненциальным ростом численности называется геометрическая прогрессия роста численности популяции. Такую способность можно наблюдать в лаборатории у бактерий, дрожжей. В не лабораторных условиях экспоненциальный рост возможно увидеть на примере саранчи или же на примере других видов насекомых. Такой рост численности вида можно наблюдать в тех местах где у него практически нет врагов, а продуктов питания более чем достаточно. В конце концов увеличение вида, после того как численность возросла в течении непродолжительного времени, рост популяции начинал снижаться.
Рассмотрим компьютерную модель размножения млекопитающих на примере модели Лотки-Вольтерры. Пусть на некоторой территории обитают два вида животных: олени и волки. Математическая модель изменения численности популяций в модели
Лотки-Вольтерры:
Начальное число жертв - xn, число хищников - yn.
Параметры модели:
P1– вероятность встречи с хищником,
P2– коэффициент роста хищников за счет жертв,
d – коэффициент смертности хищников,
a – коэффициент прироста численности жертв.
В учебной задаче были заданы такие значения: численность оленей равнялось 500, численности волков равна 10, коэффициент прироста оленей равен 0,02, коэффициент прироста численности волков равен 0,1, вероятность встречи с хищником 0,0026, коэффициент роста хищников за счет жертв 0,000056. Данные рассчитаны на 203 года.
Исследуем влияние коэффициент прироста жертв на развитие двух популяций, остальные параметры оставим без изменений.
На схеме 1 наблюдается увеличение численности жертвы и затем, с некоторым опозданием наблюдается прирост хищников. Затем хищники выбивают жертв, число жертв резко падает и вслед за ним уменьшается число хищников (рис. 1).

Рисунок 1. Численность популяций при низкой рождаемости у жертв
Проанализируем изменение модели, увеличив коэффициент рождаемости жертвы а=0,06. На схеме 2 мы видим циклический колебательный процесс, приводящий к увеличению численности обоих популяций со временем (рис. 2).
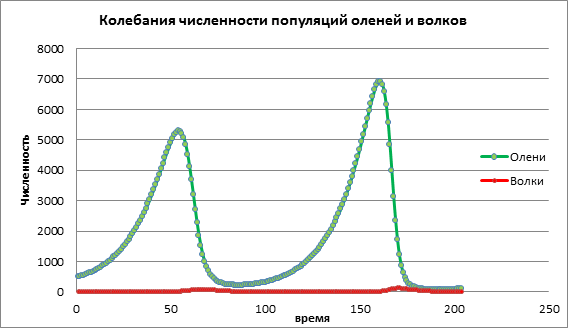
Рисунок 2.Численность популяций при средней рождаемости у жертв
Рассмотрим как изменится динамика популяций при высоком значении коэффициента рождаемости жертвы а=1,13. На рис. 3 наблюдается резкое увеличение численности обеих популяций с последующим вымиранием, как жертвы, так и хищника. Это происходит за счет того, что численность популяции жертв увеличилось до такого количества, что стали заканчиваться ресурсы, вследствие чего происходит вымирание жертвы. Вымирание хищников происходит из-за того, что сократилось количество жертв и у хищников закончились ресурсы для существования.

Рисунок 3.Численность популяций при высокой рождаемости у жертв
Исходя из анализа данных компьютерного эксперимента, можно сделать выводы о том, что компьютерное моделирование позволяет нам прогнозировать численность популяций, изучать влияние различных факторов на популяционную динамику. В приведенном примере мы исследовали модель «хищник-жертва», влияние коэффициента рождаемости жертв на численность оленей и волков. Небольшой прирост популяции жертв приводит к небольшому увеличению жертв, которую через некоторый период уничтожают хищники. Умеренный прирост популяции жертв приводит к увеличению численности обеих популяций. Высокий прирост популяции жертв приводит сначала к быстрому росту популяции жертв, это влияет на увеличение роста хищников, но затем расплодившиеся хищники быстро уничтожают популяцию оленей. В итоге оба вида вымирают.
Математическое моделирование биологических процессов началось с создания первых простейших моделей экологической системы.
Допустим, в некотором замкнутом районе живут рыси и зайцы. Рыси питаются только зайцами, а зайцы – растительной пищей, имеющейся в неограниченном количестве. Необходимо найти макроскопические характеристики, описывающие популяции. Такими характеристиками являются число особей в популяциях.
Простейшая модель взаимоотношений популяций хищника и жертвы, основанная на логистическом уравнении роста, названа (как и модель межвидовой конкуренции) по имени ее создателей - Лотки и Вольтерра. Эта модель крайне упрощает исследуемую ситуацию, но все же полезна в качестве отправной точки в анализе системы хищник-жертва.
Предположим, что (1) популяция жертвы существует в идеальной (независимой от плотности) среде, где ее рост может ограничивать только наличие хищника, (2) столь же идеальна среда, в которой существует хищник, рост популяции которого ограничивает лишь обилие жертв, (3) обе популяции размножаются непрерывно согласно экспоненциальному уравнению роста, (4) скорость поедания жертв хищниками пропорциональна частоте встреч между ними, которая, в свою очередь, является функцией плотности популяций. Эти допущения и лежат в основе модели Лотки - Вольтерра.
Пусть в отсутствие хищников популяция жертвы растет экспоненциально:
dN/dt =r 1 N 1
где N -численность, а r, - удельная мгновенная скорость роста популяции жертвы. Если же хищники присутствуют, то они уничтожают особей жертвы со скоростью, которая определяется, во-первых, частотой встреч хищников и жертв, возрастающей по мере увеличения их численностей, и, во-вторых, эффективностью, с которой хищник обнаруживает и ловит свою жертву при встрече. Число жертв, встреченных и съеденных одним хищником N с, пропорционально эффективности охоты, которую мы выразим через коэффициент С 1; численности (плотности) жертвы N и времени, затраченному на поиски Т:
N C =C 1 NT (1)
Из этого выражения легко определить удельную скорость потребления жертв хищником (т.е. число жертв, поедаемых одной особью хищника в единицу времени), которую часто называют также функциональным ответом хищника на плотность популяции жертвы:
В рассматриваемой модели С 1 является константой. Это означает, что число жертв, изъятых хищниками из популяции, линейно возрастает с увеличением ее плотности (так называемый функциональный ответ типа 1). Ясно, что общая скорость поедания жертв всеми особями хищника составит:
![]() (3)
(3)
где Р - численность популяции хищника. Теперь мы можем записать уравнение роста популяции жертвы следующим образом:
При отсутствии жертвы особи хищника голодают и гибнут. Предположим также, что в этом случае численность популяции хищника будет уменьшаться экспоненциально согласно уравнению:
![]() (5)
(5)
где r 2 - удельная мгновенная смертность в популяции хищника.
Если жертвы присутствуют, то те особи хищника, которые смогут их найти и съесть, будут размножаться. Рождаемость в популяции хищника в данной модели зависит только от двух обстоятельств: скорости потребления жертв хищником и эффективности, с которой поглощенная пища перерабатывается хищником в его потомство. Если мы выразим эту эффективность через коэффициент s, то рождаемость составит:
![]()
Поскольку С 1 и s - константы, их произведение - это также константа, которую мы обозначим как С 2 . Тогда скорость роста популяции хищника будет определяться балансом рождаемости и смертности в соответствии с уравнением:
![]() (6)
(6)
Уравнения 4 и 6 вместе образуют модель Лотки-Вольтерра.
Свойства этой модели мы можем исследовать точно так же, как и в случае конкуренции, т.е. построив фазовую диаграмму, на которой численность жертвы отложена по оси ординат, а хищника - по оси абсцисс, и проведя на ней изоклины-линии, соответствующие постоянной численности популяций. С помощью таких изоклин определяют поведение взаимодействующих популяций хищника и жертвы.
Для популяции жертвы: при откуда
Таким образом, поскольку r, и С 1 , - константы, изоклиной для жертвы будет линия, на которой численность хищника (Р) является постоянной, т.е. параллельная оси абсцисс и пересекающая ось ординат в точке Р =r 1 / С 1 . Выше этой линии численность жертвы будет уменьшаться, а ниже- возрастать.
Для популяции хищника:
при откуда
Поскольку r 2 и С 2 - константы, изоклиной для хищника будет линия, на которой численность жертвы (N) является постоянной, т.е. перпендикулярная оси ординат и пересекающая ось абсцисс в точке N = r 2 /С 2 . Слева от нее численность хищника будет уменьшаться, а справа - возрастать.
Если мы рассмотрим эти две изоклины вместе, то легко заметим, что взаимодействие популяций хищника и жертвы имеет циклический характер, поскольку их численности претерпевают неограниченные сопряженные колебания. Когда велико число жертв, численность хищников растет, что приводит к увеличению пресса хищничества на популяцию жертвы и тем самым к снижению ее численности. Это снижение, в свою очередь, ведет к нехватке пищи у хищников и падению их численности, которое вызывает ослабление пресса хищничества и увеличению численности жертвы, что снова приводит к росту популяции жертвы и т.д.
Для данной модели характерна так называемая "нейтральная стабильность", которая означает, что популяции неограниченно долго совершают один и тот же цикл колебаний до тех пор, пока какое-либо внешнее воздействие не изменит их численность, после чего популяции совершают новый цикл колебаний с иными параметрами. Для того, чтобы циклы стали стабильными, популяции должны после внешнего воздействия стремиться вернуться к первоначальному циклу. Такие циклы, в отличие от нейтрально стабильных колебаний в модели Лотки-Вольтерра, принято называть устойчивыми предельными циклами.
Модель Лотки-Вольтерра, тем не менее, полезна тем, что позволяет продемонстрировать основную тенденцию в отношениях хищник-жертва возникновение циклических сопряженных колебаний численности их популяций.
к договору от ___.___,20___ г. об оказании платных образовательных услуг
Министерство образования и науки Российской Федерации
Лысьвенский филиал
Пермского Государственного технического университета
Кафедра ЕН
по дисциплине «Моделирование систем»
тема: Система хищник-жертва
Выполнил:
Студент гр. БИВТ-06
------------------
Проверил преподаватель:
Шестаков А. П.
Лысьва, 2010 г.
Реферат
Хищничество - трофические отношения между организмами, при которых один из них (хищник) атакует другого (жертву) и питается частями его тела, то есть обычно присутствует акт умерщвления жертвы. Хищничество противопоставляется поеданию трупов (некрофагии) и органических продуктов их разложения (детритофагии).
Довольно популярно также другое определение хищничества, предлагающее хищниками называть лишь организмы, поедающие животных, в отличие от растительноядных, поедающих растения.
Кроме многоклеточных животных, в роли хищников могут выступать протисты, грибы и высшие растения.
Размер популяции хищников влияет на размер популяции их жертв и наоборот, динамика популяций описывается математической моделью Лотки-Вольтерра, однако данная модель является высокой степенью абстракции, и не описывает реальные взаимосвязи между хищником и жертвой, и может рассматриваться только как первая степень приближения математической абстракции.
В процессе совместной эволюции хищники и жертвы приспосабливаются друг к другу. У хищников появляются и развиваются средства обнаружения и атаки, а у жертв - средства скрытности и защиты. Поэтому наибольший вред жертвам могут нанести новые для них хищники, с которыми те не вступали ещё в «гонку вооружений».
Хищники могут специализироваться на одном–нескольких видах для добычи, это делает их в среднем более успешными в охоте, но повышает зависимость от данных видов.
Система хищник-жертва.
Взаимодействие хищник-жертва - основной тип вертикальных взаимоотношений организмов, при котором по пищевым цепям передаются вещество и энергия.
Равновесность В. х. - ж. наиболее легко достигается в том случае, если в пищевой цепи имеется не менее трех звеньев (например, трава - полёвка - лисица). При этом плотность популяции фитофага регулируется взаимоотношениями как с нижним, так и с верхним звеном пищевой цепи.
В зависимости от характера жертв и типа хищника (истинный, пастбищник) возможна разная зависимость динамики их популяций. При этом картина осложняется тем, что хищники очень редко бывают монофагами (т. е. питающимися одним видом жертвы). Чаще всего, когда истощается популяция одного вида жертвы и ее добывание требует слишком больших затрат сил, хищники переключаются на другие виды жертв. Кроме того, одну популяцию жертв может эксплуатировать несколько видов хищников.
По этой причине часто описываемый в экологической литературе эффект пульсирования численности популяции жертвы, за которым с определенным запаздыванием пульсирует численность популяции хищника, в природе встречается крайне редко.
Равновесие между хищниками и жертвами у животных поддерживается специальными механизмами, исключающими полное истребление жертв. Так, жертвы могут:
- убегать от хищника (в этом случае в результате соревнования повышается подвижность и жертв, и хищников, что особенно характерно для степных животных, которым негде прятаться от преследователей);
- приобретать защитную окраску (<притворяться> листьями или сучками) или, напротив, яркий (например, красный) цвет, предупреждающий хищника о горьком вкусе;
- прятаться в укрытия;
- переходить к мерам активной обороны (рогатые травоядные, колючие рыбы), часто совместной (птицы-жертвы коллективно отгоняют коршуна, самцы оленей и сайгаков занимают <круговую оборону> от волков и т. д.).
система РА88, которая одновременно предсказывает вероятность более чем 100 фармакологических эффектов и механизмов действия вещества на основе его структурной формулы. Эффективность применения этого подхода к планированию скрининга составляет около 800%, а точность компьютерного прогноза на 300% превосходит предсказание экспертов.
Итак, одним из конструктивных инструментов получения новых знаний и решений в медицине является метод математического моделирования. Процесс математизации медицины – частое проявление взаимопроникновения научных знаний, повышающее эффективность лечебно-профилактической работы.
4. Математическая модель «хищники-жертвы»
Впервые в биологии математическую модель периодического изменения числа антагонистических видов животных предложил итальянский математик В. Вольтерра с сотрудниками. Модель, предложенная Вольтерра, явилась развитием идеи, намеченной в 1924 году А. Лоттки в книге "Элементы физической биологии". Поэтому эта классическая математическая модель известна как модель "Лоттки-Вольтерра".
Хотя в природе отношения антагонистических видов более сложные, чем в модели, тем не менее они являются хорошей учебной моделью, на которой можно изучать основные идеи математического моделирования.
Итак, задача : в некотором экологически замкнутом районе живут два вида животных (например, рыси и зайцы). Зайцы (жертвы) питаются растительной пищей, имеющейся всегда в достаточном количестве (в рамках данной модели не учитывается ограниченность ресурсов растительной пищи). Рыси (хищники) могут питаться только зайцами. Необходимо определить, как будет меняться численность жертв и хищников с течением времени в такой экологической системе. Если популяция жертв увеличивается, вероятность встреч хищников с жертвами возрастает, и, соответственно, после некоторой временной задержки, растет популяция хищников. Эта достаточно простая модель вполне адекватно описывает взаимодействие между реальными популяциями хищников и жертв в природе.
Теперь приступим к составлению дифференциальных уравнений. Обо-
значим число жертв через N, а число хищников через M. Числа N и M являются функциями времени t . В нашей модели учтем следующие факторы:
а) естественное размножение жертв; б) естественная гибель жертв;
в) уничтожение жертв за счет поедания их хищниками; г) естественное вымирание хищников;
д) увеличение числа хищников за счет размножения при наличии пищи.
Так как речь идет о математической модели, то задачей является получение уравнений, в которые входили бы все намеченные факторы и которые описывали бы динамику, то есть изменение числа хищников и жертв со временем.
Пусть за некоторое время t количество жертв и хищников изменится на ∆N и ∆M. Изменение числа жертв ∆N за время ∆t определяется, во-первых, увеличением в результате естественного размножения (которое пропорционально имеющемуся количеству жертв):
где В – коэффициент пропорциональности, характеризующий скорость естественного вымирания жертв.
В основе вывода уравнения, описывающего уменьшение числа жертв изза поедания их хищниками, лежит идея о том, что чем чаще происходит их встреча, тем быстрее уменьшается число жертв. Ясно также, что частота встреч хищников с жертвой пропорционально и числу жертв и числу хищников, то
Поделив левую и правую часть уравнения (4) на ∆t и перейдя к пределу при ∆t→0 , получим дифференциальное уравнение первого порядка:
Для того, чтобы решить это уравнение, нужно знать, как меняется число хищников (М ) со временем. Изменение числа хищников (∆М ) определяется увеличением из-за естественного размножения при наличии достаточного количества пищи (М 1 = Q∙N∙M∙∆t ) и уменьшением из-за естественного вымирания хищников (M 2 = - P∙M∙∆t ):
M = Q∙N∙M∙∆t - P∙M∙∆t |
Из уравнения (6) можно получить дифференциальное уравнение:
Дифференциальные уравнения (5) и (7) представляют собой математическую модель "хищники-жертвы". Достаточно определить значения коэффици-

ентов A, B, C, Q, P и математическую модель можно использовать для решения поставленной задачи.
Проверка и корректировка математической модели. В данной лабора-
торной работе предлагается кроме просчета наиболее полной математической модели (уравнения 5 и 7), исследовать более простые, в которых что-либо не учитывается.
Рассмотрев пять уровней сложности математической модели, можно "почувствовать" этап проверки и корректировки модели.
1-ый уровень – в модели учтено для "жертв" только их естественное размножение, "хищники" отсутствуют;
2-ой уровень – в модели учтено для "жертв" их естественное вымирание, "хищники" отсутствуют;
3-ий уровень – в модели учтены для "жертв" их естественное размножение
и вымирание, "хищники" отсутствуют;
4-ый уровень – в модели учтены для "жертв" их естественное размножение
и вымирание, а также поедание "хищниками", но число "хищников" остается неизменным;
5-ый уровень – в модели учтены все обсуждаемые факторы.
Итак, имеем следующую систему дифференциальных уравнений:
где М – число "хищников"; N – число "жертв";
t – текущее время;
A – скорость размножения "жертв"; C – частота встреч "хищники-жертвы"; B – скорость вымирания "жертв";
Q – размножение "хищников";
P – вымирание "хищников".
1-ый уровень: М = 0, В = 0; 2-ой уровень: М = 0, А = 0; 3-ий уровень: М = 0; 4-ый уровень: Q = 0, Р = 0;
5-ый уровень: полная система уравнений.
Подставляя значения коэффициентов в каждый уровень, будем получать разные решения, например:
Для 3-его уровня значение коэффициента М=0 , тогда

решая уравнение получим
Аналогично для 1-го и 2-го уровней. Что касается 4-го и 5-го уровней, то здесь необходимо решать систему уравнений методом Рунге-Кутта. В результате получим решение математических моделей данных уровней.
II. РАБОТА СТУДЕНТОВ ВО ВРЕМЯ ПРАКТИЧЕСКОГО ЗАНЯТИЯ
Задание 1 . Устно-речевой контроль и коррекция усвоения теоретического материала занятия. Сдача допуска к занятию.
Задание 2 . Выполнение лабораторной работы, обсуждение полученных результатов, оформление конспекта.
Выполнение работы
1. С рабочего стола компьютера вызвать программу "Лаб. №6", щелкнув по соответствующему ярлыку два раза левой клавишей "мыши".
2. Щелкнуть дважды левой клавишей "мыши" по ярлыку "PREDATOR".
3. Выбрать ярлык "PRED" и повторить вызов программы левой клавишей "мыши" (щелкнув дважды).
4. После титульной заставки нажать "ENTER".
5. Моделирование начинать с 1-го уровня.
6. Ввести год, с которого будет проводиться анализ модели: например, 2000
7. Выбрать временные интервалы, например, в течение 40 лет, через 1 год (затем через 4 года).
2-ой уровень: B = 0.05; N0 = 200;
3-ий уровень: A = 0.02; B = 0.05; N = 200;
4-ый уровень: A = 0.01; B = 0.002; C = 0.01; N0 = 200; M = 40; 5-ый уровень: A = 1; B = 0.5; C = 0.02; Q = 0.002; P = 0.3; N0 = 200;
9. Подготовить письменный отчет по работе, который должен содержать уравнения, графики, результаты расчета характеристик модели, выводы по проделанный работе.
Задание 3. Контроль конечного уровня знаний:
а) устно-речевой отчет за выполненную лабораторную работу; б) решение ситуационных задач; в) компьютерное тестирование.
Задание 4. Задание на следующее занятие: раздел и тема занятия, согласование тем реферативных докладов (объем доклада 2-3 стр., регламент 5-7 мин.).

 Замок из костей в праге. Храм из костей. Церковь из костей
Замок из костей в праге. Храм из костей. Церковь из костей Классификация ценных бумаг Классификация и виды ценных бумаг
Классификация ценных бумаг Классификация и виды ценных бумаг Как сделать магический маятник в домашних условиях
Как сделать магический маятник в домашних условиях Смотреть что такое "Хаксли, Джулиан" в других словарях Отрывок, характеризующий Хаксли, Джулиан
Смотреть что такое "Хаксли, Джулиан" в других словарях Отрывок, характеризующий Хаксли, Джулиан